Den 2. ATP-syntasen -redder planeten
Av David Coppedge 10.juli 2023; oversatt herfra
ATP -syntase har blitt omtalt her mange ganger på grunn av sin utsøkte rotasjonsmekanisme og effektive drift. Seere av animasjoner som vår på YouTube -lenke trenger vanligvis lite overbevisende argumenter for at den ser designet ut. Og når de lærer flere detaljer, som 6000 o / min hastighet, dens veivaksel og tredelt ATP-produksjonssenter, blir intuisjonen vanskelig å fjerne, selv når evolusjonister insisterer på at det dukket opp ved en tilfeldighet. Det er et velfortjent ikon for intelligent design.
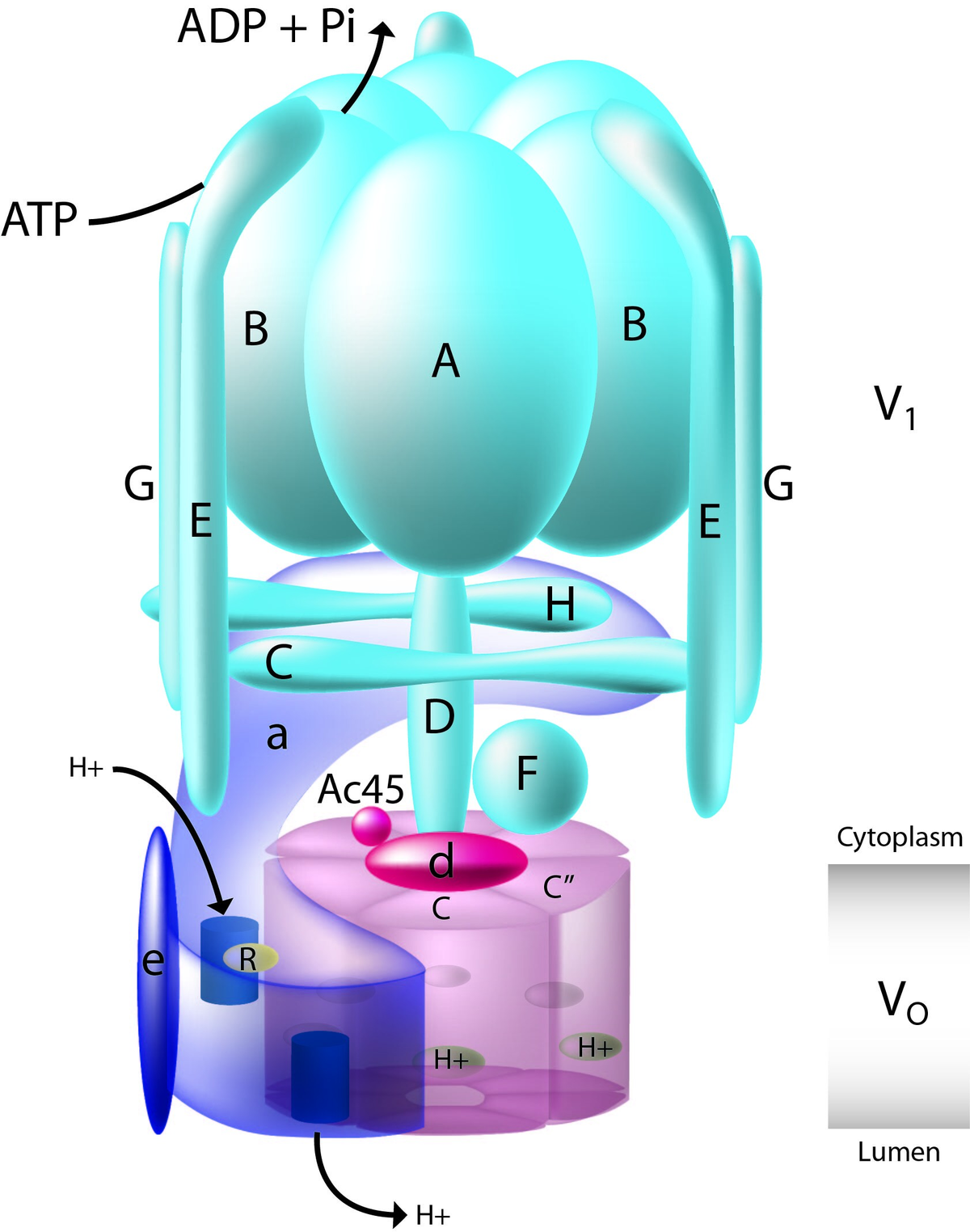 Bilde 1. V-ATPase- fra wikipedia
Bilde 1. V-ATPase- fra wikipedia
Den som oftest refereres til er F-typen av ATP-syntase vist i videoen. Men det er en annen-ATPase av V-typen-det er ikke mindre underlig. Det ser ut som F-typen, men V-type ATPases (la oss kalle dem VHA) fungerer omvendt: I stedet for å bruke protonmotivkraften generert av elektrontransportkjeden for å produsere ATP, bruker de ATP "energivaluta" -molekyler til å pumpe protoner inn i organeller, og dermed øke surheten. De finnes ofte på membranene til vakuoler som trenger H+ -ioner for å senke pH for fagocytose eller andre typer fordøyelses- eller resirkuleringsfunksjoner. Forskere har nå vist at de er mer distribuert i cellen enn antatt.
Kroppscellene våre inneholder begge typer ATPase. Men hvorfor sier jeg at disse VHA -rotasjonsmotorene redder planeten? En rapport fra Daniel P. Yee in Current Biology forteller historien -lenke.
Det begynner i havet
Diatomer, dinoflagellater og kokolitoforer er dominerende grupper av marin eukaryotisk planteplankton, som samlet er ansvarlige for mesteparten av primærproduksjonen i havet.
Primærproduksjon representerer bunnen av næringskjeden, som høyere organismer er avhengige av. Fotosyntetiske mikrober i havet er de største aktørene. For nåværende formål kan vi ignorere forfatterenes evolusjonshistorie om hvordan disse marine mikroberne oppnådde VHA -ene med en eller annen "selektiv fordel." Det er nok å fokusere på hva molekylære maskiner gjør:
"Siden intracellulære fordøyelsesvakuoler allestedsnærværende blir forsuret, ved V-type H+-Atpase (VHA), ble protonpumper foreslått for å forsurte mikro-miljøet rundt sekundære kloroplaster for å fremme dehydrasjonen av oppløst inorganisk karbon (OIK) til CO2, og dermed en forbedring av oppløst inorganisk karbon til CO2, og slik forbedre fototsyntesen.
Vi rapporterer at VHA er lokalisert rundt kloroplaster av sentriske diatomer og at VHA bidrar betydelig til deres fotosyntese over et bredt spekter av oseaniske bestråling."
Mer fotosyntese betyr mer primærproduksjon, og større støtte for en mangfoldig biosfære.
Bilde 2. Subcellulaer-lokalisering-av-VHA-i-T-pseudonana
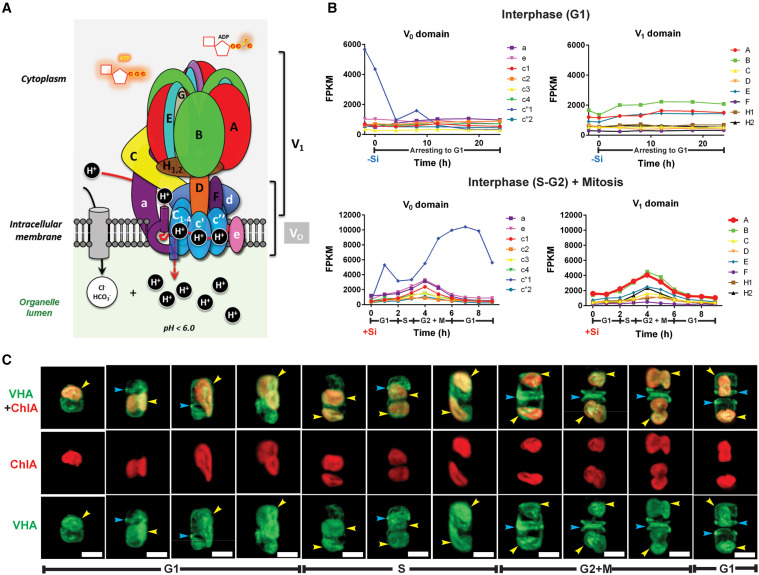 "Basert på bidrag fra diatomer til biogeokjemiske sykluser, bidrar VHA-mediert forbedring av fotosyntesen minst 3,5 GigaTonn med fast karbon per år (eller 7% av primærproduksjonen i havet), og gir et eksempel på en symbiose-avledet, evolusjonær innovasjon med Globale miljømessige implikasjoner."
"Basert på bidrag fra diatomer til biogeokjemiske sykluser, bidrar VHA-mediert forbedring av fotosyntesen minst 3,5 GigaTonn med fast karbon per år (eller 7% av primærproduksjonen i havet), og gir et eksempel på en symbiose-avledet, evolusjonær innovasjon med Globale miljømessige implikasjoner."
Nok en gang er den evolusjonære historien ikke-essensiell for den overraskende implikasjonen: alle livsfordeler, ved forbedring av fotosyntesen levert av VHA-molekylmotorer i diatomer. Antallet på 7 prosent er utvilsomt mye høyere, hvis de andre dominerende gruppene av marin planteplankton er innarbeidet i. Ytterligere tester viste forskerteamet som lignende VA-mediert forbedring av fotosyntesen, forekommer i kokolitoforer, dinoflagellater og sannsynligvis i alle fotosyntetiske organismer.
Artikkelens diagram over VHA i figur 2 ser nesten identisk ut med F-typen ATP-syntase bortsett fra retningen på proton (H+ ion) -strømmen. Begge typene kan beskrives av ID -talsmenn som irreduserbare, komplekse molekylære motorer basert på delelisten alene:
"VHA er et holoenzymproteinkompleks som er sammensatt av 16 underenheter, med en membran som spenner over V0-domene og cytosolisk vendt katalytisk V1-domene (figur 2A). Transkriptomisk analyse av synkroniserte kulturer av T. pseudonana [en sentrisk diatom] demonstrerte konstitutivt mRNA -ekspresjon av alle VHA -underenheter, noe som antydet at VHA, er viktig i hele cellesyklusen (figur 2B; lenke). Imidlertid kan VHA -holoenzym ha flere subcellulære lokaliseringer og funksjoner."
Mens den kjente F-typen ATP-syntase er lokalisert til mitokondriene (hos dyr) eller kloroplaster (i fotosyntetiske mikrober og planter), finnes V-typer på andre subcellulære steder. De utfører flere funksjoner på grunn av sin proton-pumpende handling, som kan justere pH i omgivelsene. Dette har gjort dem vanskelige å studere. Noen få funksjoner er kjent.
Men hvor mange andre er det?
"I diatomer er det rapportert VHA i membranene til vakuoler, kloroplast endoplasmatisk retikulum (CER) og silikaavsetningsvesikler (SDV), hvor det strengt tatt er nødvendig for biomineralisering av silisiumscellevegg og celledeling. Kompleksiteten og funksjonell allsidighet av VHA er utfordringer for genetiske manipulasjonsmetoder, som konstitutivt destabiliserer flere fysiologiske funksjoner og forvirrende fenotypisk tolkning. De utelukker også bruk av transkriptomikk for å utlede den fysiologiske rollen til VHA, da disse analysene ikke kan gi informasjon om den subcellulære lokaliseringen av VHA -holoenzymet."
Forfatterne var begeistret for en annen implikasjon av oppdagelsen deres: karbonsekvestrering. Fordi VHA-maskiner forbedrer fotosyntesen betydelig, betyr det at mer karbon blir omdannet til CO2 som kloroplasten trenger for sukkersyntese. Den svake pH -reduksjonen gitt av VHA -protonpumpene endrer kjemien for å favorisere CO2 -produksjonen, noe som ville være dårlig; Men samtidig skaper det en gradient som favoriserer å flytte CO2 inn i kloroplasten i stedet for inn i cytoplasmaet, der den ville rømme ut i atmosfæren. Som et resultat frigjøres mer oksygen ved fotosyntese, mindre CO2 frigjøres ut i atmosfæren under fotosyntesen, og når kiselalgen dør og faller til havbunnen, tar de det faste karbonet med seg. V-type ATPases utfører således en annen global funksjon for biosfæren: en karbonkonsentrerende mekanisme.
"VHA er et universelt trekk ved eukaryote celler og er til stede i en rekke organeller, inkludert endosomer, fagosomer, makropinosomer, lysosomer, golgi og melanosomer. Siden VHA alltid forsurer lumen til hver av disse organellene til pH 6, slutter vi ut at VHA i CER/PP -membranene til T. pseudonana må oppnå en lignende effekt. Dette er sentralt, fordi ved pH 6,3 ekvilibrerer flertallet av OIK (Oppløst Inorganisk Karbon) til CO2. Derfor foreslår vi at VHA fremmer CO2 -akkumulering i mikro -miljøet utenfor kloroplasten. I samsvar med modellsimuleringer av OIK -flukser i diatom CCM, ville noe av CO2 diffundere tilbake til cytoplasma (pH
6, slutter vi ut at VHA i CER/PP -membranene til T. pseudonana må oppnå en lignende effekt. Dette er sentralt, fordi ved pH 6,3 ekvilibrerer flertallet av OIK (Oppløst Inorganisk Karbon) til CO2. Derfor foreslår vi at VHA fremmer CO2 -akkumulering i mikro -miljøet utenfor kloroplasten. I samsvar med modellsimuleringer av OIK -flukser i diatom CCM, ville noe av CO2 diffundere tilbake til cytoplasma (pH  7,2). Imidlertid etablerer den høyere pH i kloroplaststroma (pH 8,15) en gunstigere delvis trykkgradient for CO2 -diffusjon i dette rommet (figur S3B). De neste trinnene følger den etablerte CCM for mikroalger. I stroma hydrerte CO2IS umiddelbart inn i HCO3- under katalyse med karbonanhydrase, som blir skutt inn i thylakoidlumen der pH er 6 på grunn av H+ pumping assosiert med fotosyntetisk elektrontransportkjede. Ved den sure pH, hCO3- dehydrater i CO2
7,2). Imidlertid etablerer den høyere pH i kloroplaststroma (pH 8,15) en gunstigere delvis trykkgradient for CO2 -diffusjon i dette rommet (figur S3B). De neste trinnene følger den etablerte CCM for mikroalger. I stroma hydrerte CO2IS umiddelbart inn i HCO3- under katalyse med karbonanhydrase, som blir skutt inn i thylakoidlumen der pH er 6 på grunn av H+ pumping assosiert med fotosyntetisk elektrontransportkjede. Ved den sure pH, hCO3- dehydrater i CO2 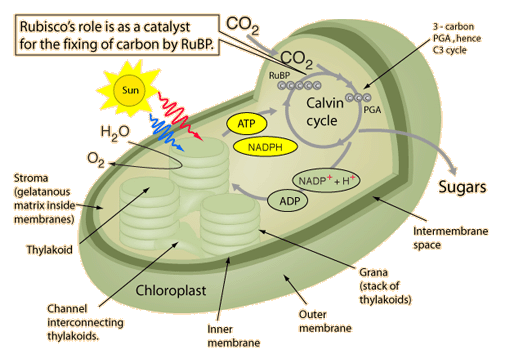 og diffunderer inn i pyrenoid-matrisen, gjennomsyrer/metter Rubisco for å maksimere karbonfikseringshastigheter."
og diffunderer inn i pyrenoid-matrisen, gjennomsyrer/metter Rubisco for å maksimere karbonfikseringshastigheter."
En bemerkelsesverdig synergi
Dette viser en bemerkelsesverdig synergi av molekylære maskiner og kjemi i et sterkt lokalisert mikro -miljø. VHAS surifiserer kloroplastens ytre, og hjelper karbonanhydrase til å bruke overflødig CO2 for å produsere flere bikarbonat-ioner. PH-gradienten favoriserer disse negative ionene til å strømme mot protonmotivkraften, som genereres av elektrontransportkjeden (komplekser I-IV) som driver F-typen ATP-syntasemotorer (kompleks V). Når bikarbonat-ionene dehydrerer tilbake til CO2 der pH synker inne i kloroplasten, metter de Rubisco-enzymer, som omdanner CO2 til næringsstoffer for livet; lenke.
Legg merke til denne nøye pH-medierte serien med delikate kjemiske reaksjoner. Innenfor dette mikro-miljøet blir CO2 brukt der det er nødvendig, men frigjøres ikke i atmosfæren. Produktene fra Rubisco favoriserer karbonforbindelser som vil bli begravet i havbunnen. Resultatet? En karbonkonsentrerende mekanisme som hjelper atmosfæren med å få mer oksygen, men mindre karbondioksid, samtidig som den beriker biosfæren med næringsstoffer. Vha, den "andre" ATP -syntasen, reddes planeten! VHA bør sees på som en nøkkelaktør i karbonsyklusen, oksygensyklusen, næringskjeden og klimaet.
Bilde 3. Rubisco enzymets viktige rolle i kretsløpet
Nesten uhemmet undring
Forfatterne kan knapt beherske undringen sin i alt dette:
"Oppsummert demonstrerte O2-produksjon, 13C-nanosims og 14C-P-E-målinger at VHA bidrar betydelig til fotosyntetisk karbonfiksering, som beholdes som biomasse. Gitt at diatomer bidrar med nesten 50% av karbonfiksering i havet, representerer komponenten i fotosyntesen som avhenger av VHA mellom 7% og 25% av oseanisk primærproduksjon, eller mellom 3,5 og 13,5 gtons fast karbon per år (tabell S3) . Disse tallene kan bare øke etter regnskap for VHA-mediert fotosyntese i andre sekundære endosymbiotiske planteplanktoner (figur 1) og fotosymbiotiske virvelløse dyr. I tillegg utgjør diatomer omtrent halvparten av biomassen som synker ned i havets indre. Basert på våre målinger av bidraget fra VHA, både på brutto og netto produktivitet i diatomer, er VHA-mediert karbonfiksering rede til å bidra betydelig til den biologiske pumpen, som skifter organisk karbon til havets interiør, og -på en geologisk tidsskala, til biomasse Begravet i den kontinentale marginen som dannet fossile drivstoffavsetninger. Selv ved det mest konservative estimatet, er samhandlingen av VHA for forbedring av fotosyntesen, en symbiose-avledet evolusjonær innovasjon med globale miljømessige implikasjoner."
En dag i fremtiden, når biologer endelig innrømmer at evolusjonen ikke er i stand til innovasjon av komplekse, fint justerte samarbeidssystemer, vil alle forskere undre seg over den intelligente designen som gir oss luft til å puste og spiselig karbon å innta, under et moderat klima. Noen av oss er foran det skjemaet.

Om forfatteren: DAVID COPPEDGE -kredit til
David Coppedge (Bilde 4)
David Coppedge er en frilans vitenskapsreporter i Sør-California. Han har vært styremedlem i Illustra Media siden grunnleggelsen og fungerer som deres vitenskapskonsulent. Han jobbet ved NASAs Jet Propulsion Laboratory (JPL) i 14 år, på Cassini-oppdraget til Saturn, til han ble kastet ut i 2011 for å dele materiale om intelligent design, en diskriminerende handling som førte til en nasjonalt publisert rettssak i 2012. Discovery Institute støttet saken hans, men en ensom dommer dømte ham mot ham, uten forklaring. En naturfotograf, friluftsmann og musiker, David har B.S. grader i realfagsutdanning og i fysikk og holder presentasjoner om ID og andre vitenskapelige emner.
Oversettelse og bilder ved Asbjørn E. Lund